



Wnt signaling pathway diagram
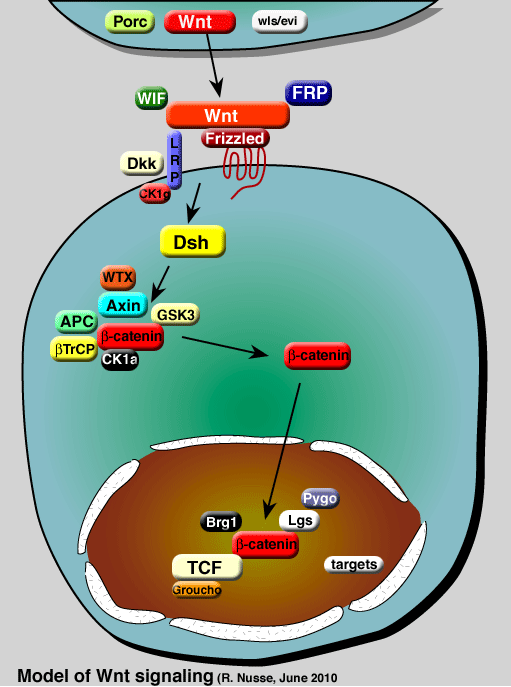
The Wnt pathway is increasingly becoming more complex and new participants are still being uncovered. It becomes hard to include all of these new components and the list below is selective. See the simplified pathway map for a further selection.
Wnt, Porc, Wls/Evi |
|---|
|
Wnt genes can be expressed in many different cell types; their expression is not restricted to dedicated cells. There is control over the secretion and processing of the Wnt protein. Wnt proteins are modified by palmitoylation (Willert 2003) and glycosylation (Mason, 1992). A special form of monounsaturated palmitoylation has been detected on a serine residue in the Wnt protein (Takada 2006) The Porcupine (Porc) protein may be involved in secretion or ER transport, as Wingless is retained in the ER in porcupine mutant Drosophila embryos (Kadowaki 1996, van den Heuvel 1993). In C. elegans, the porcupine homolog mom-1 has a similar function in promoting secretion of the Wnt protein Mom-2 (Rocheleau 1997). Porcupine has some homology to a family of o-acyl transferases and may be involved in lipid modification of Wnt proteins (Hofmann, 2000, Willert 2003, Zhai, 2004, Takada 2006). In addition to porcupine, several other proteins, including the transmembrane Wls/Evi are specifically involved in Wnt secretion (Banziger, 2006; Bartscherer, 2006). Even though there is control over Wnt secretion, there is no evidence that the function of genes like porcupine and Wls/Evi is restricted to particular cells implying that once Wnt genes are expressed, their proteins will be made in any cell type. |
Frp, WIF, Dkk |
|---|
|
In the extracellular space, several secreted proteins can bind directly to Wnts, to modulate Wnt activity. The secreted Frps (Rattner, 1997) resemble the ligand binding domains of the Frizzled receptor. WIFs form another group of secreted Wnt binding factors (Hsieh 1999). Dickkopf (Dkk) in Xenopus antagonizes Wnt action (Glinka1998 Fedi, 1999) by binding to LRP (Mao et al,2001, Bafico et al, 2001; Semenov et al, 2001). |
Frizzled, LRP/Arrow, Dsh |
|---|
|
Wnts interact genetically and biochemically with a complex of receptors.The specificity between Wnts and receptor complexes is determined by the Frizzled class of receptors (Bhanot, 1996), of which the CRD (cysteine-rich domain) is the primary ligand binding domain (Dann et al, 2001). In Drosophila as well as in vertebrates, LRP (or arrow) is required for Wnt signaling as well, and can bind to Wnt-Frizzled to form a ternary complex (Wehrli et al, 2000: Tamai et al, 2000; Pinson et al, 2000). The cytoplasmic tail of LRP can bind to Axin, in a Wnt and phosphorylation dependent manner (Mao et al., 2001, Tolwinsky, 2003, Tamai et al, 2004). Phosphorylation of the tail of LRP is regulated by two protein kinases: GSK3 and CK1gamma (Zeng, 2005; Davidson 2005; reviewed by Nusse, 2005 . Zeng et al (2008) propose a role for Dsh and Fz in this process as well. The Wnt signal leads, through its receptor to activation of Dishevelled (Dsh). Wnt signaling at the level of receptor interactions and activity may be associated with intracellular compartments such as vacuoles, given the role of the pro-renin receptor as an adaptor between Wnt receptors and the vacuolar H+-adenosine triphosphatase (V-ATPase) complex (Cruciat, 2010). |
beta-catenin , GSK3, Axin, WTX, APC, beta-TrCP |
|---|
|
Armadillo/beta-catenin is the key mediator of the Wnt signal. In cells not exposed to the signal, beta-catenin levels are kept low through interactions with the protein kinase GSK-3b, CK1a, APC and Axin (Behrens, 1998 Itoh 1998., Hamada, 1999.) Another player in this complex is the Wilms tumor suppressor gene WTX (Major, 2007, Rivera, 2007) beta-catenin is degraded, after phosphorylation by GSK-3 and CK1 alpha (Yanagawa 2002, Liu 2002, Amit 2002), through the ubiquitin pathway (Aberle 1997.), involving interactions with beta-TrCP(Jiang 1998, Marikawa 1998,; reviewed in Maniatis 1999) In a current model, Wnt signaling initially leads to a complex between Dsh, GBP/Frat1, Axin and Zw3/GSK, which may be the regulatory step in the inactivation of Zw3/GSK (Salic, 2000; Farr 2000). The DIX domain in Axin is similar to the NH2 terminus in Dsh, and promotes interactions between Dsh and Axin (Hsu 1999, Smalley, 1999). As a consequence, GSK does not phosphorylatebeta-catenin anymore, releasing it from the Axin complex and accumulation (Salic, 2000).The stabilizedbeta-catenin then enters the nucleus (Tolwinski and Wieschaus, 2004) to interact with TCF. Binding of Axin to the cytoplasmic tail of LRP in a Wnt dependent manner (Mao et al., 2001, Tolwinski 2003, Tamai et al, 2004) may also play a role in rearranging this complex. Goentero and Kirschner (2009) have shown that abolute levels of beta-catenin are less important than fold-changes in determining activity of Wnt signaling in target cells (reviewed by Ferrell, 2009). Loss of APC in mammalian cells can also lead to a critical loss over beta-catenin control, leading to cell transformation (reviewed in Polakis, 2000). |
TCF, Groucho, Brg-1, Lgs, Pygo, Hyx |
|---|
|
In the nucleus, in the absence of the Wnt signal,TCF acts as a repressor of Wnt/Wg target genes (Brannon 1997, Bienz 1998 . Riese 1997; also in C. elegans Lin 1998). TCF can form a complex with Groucho (Cavallo 1998).beta-catenin can convert TCF into a transcriptional activator of the same genes that are repressed by TCF alone (reviewed in Nusse, 1999). Daniels and Weis (2005) have shown that beta-catenin displaces Groucho from TCF. Two other key players in this complex are Legless (Bcl9) and Pygopos (Kramps 2002, Thompson 2002, Parker 2002). Brg-1 is a mammalian SWI/SNF and Rsc chromatin-remodelling complex protein binding tobeta-catenin and promoting activity (Barker, 2001) There are many target genes of the Wnt pathway, listed in a separate table. |