| Cognitive Neuroscience: Notes
for Wine Lecture
GENES AND BEHAVIOR ÓJ. J. Wine, 2000 [Last edited 3-28-2000]
Does human behavior have a genetic component? The answer to that question is controversial in part because ethical and legal issues make controlled studies of human behavior difficult to devise. However, the evidence for a genetic component in human behavior is overwhelming in spite of that limitation.
All behavior is the joint product of heredity and environment, but differences in behavior can be apportioned between hereditary and environment. The Canadian psychologist Donald Hebb has likened the nature-nurture controversy to an argument about whether the area of a rectangle depends more importantly on its width or length. For any given rectangle the area is always a joint product of the two dimensions. However, when comparing two rectangles having different areas, it is meaningful to ask to what extent the different areas can be attributed to differences in either of the dimensions. (Note the corollary: two rectangles can have the same area but different dimensions). Substituting, we can see that any behavior is always the joint product of heredity and environment, but differences in behavior can be apportioned between differences in heredity and in environment. Having said that, it is extremely difficult to go from genes to behavior, or more generally to bridge the chasm between genotype and phenotype. A single genes usually makes a single protein, or sometimes only a part of a protein (for example, it takes the products of 4 different genes to produce a single acetylcholine receptor/channel). A typical cell expresses ~10,000 different gene products. Therefore, if the product of a single gene differs from the prototype for that gene because of a heritable change in the gene, we would expect the following: (Why do you think most mutations are deleterious?) The human retina contains receptors that transduce light into electrical signals. The signals are then relayed via synapses to other neurons and ultimately to the brain. Color vision is made possible by the cone receptors. You will hear a great deal about color vision in later lectures. For a simple, online introduction, see Breaking the Code of Color: Unlike altered or lost color vision, in which the pathway from gene to behavior is well understood, Huntingtons disease remains an enigma even though the gene was discovered in 1993. Huntington's disease (or Huntington's chorea, from the dance-like movements made by some patients) is a dominant disease: on average it affects half of all members of a family in which one parent is affected. It is passed on because individuals who carry the gene usually do not realize they are affected until middle age, although onset of symptoms can vary by more than 50 years. Both symptoms and post-mortem examination of brains indicate that brain damage occurs most severely in the basal ganglia, thus first affecting motor behavior but eventually affecting cognition and death. Even though a single gene is involved, the age of onset and time course of the disease varies greatly. About 30,000 Americans have HD. Dominant diseases usually involve a gain of function, rather than the mere loss of a protein. Indeed, the discovery of the gene for Huntington's disease, after a long and much publicized search, shows that the disease results from the expansion of a tract of repeated CAG nucleotides at the beginning of the coding region of the gene. In normal individuals this tract is 6-39 triplets in length, but people with Huntingtons disease have what is called a triplet expansion in which the tract length increases up to 180. The age of onset decreases as the tract length increases: so lengths of 40-55 make up the majority of adult onset cases, while expansions above 70 cause juvenile onset. The triplet CAG codes for the amino acid glutamine, 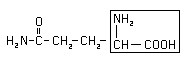 so
the triplet expansion means that the protein will have a longer than normal
tract of glutamine at its N terminus. The protein coded by the Huntingtons
disease gene (i.e. by the gene that causes Huntingtons disease when it
is mutated) is called huntingtin.
It is a giant protein of 350 kDa, and, surprisingly, it is found in virtually
every type of cell examined. Huntingtin is an essential protein--when
it is eliminated in mice by gene targeting, the mice die as embryos.
Is function is unknown. Does Huntingtons disease provide a clue
to the function of huntingtin? so
the triplet expansion means that the protein will have a longer than normal
tract of glutamine at its N terminus. The protein coded by the Huntingtons
disease gene (i.e. by the gene that causes Huntingtons disease when it
is mutated) is called huntingtin.
It is a giant protein of 350 kDa, and, surprisingly, it is found in virtually
every type of cell examined. Huntingtin is an essential protein--when
it is eliminated in mice by gene targeting, the mice die as embryos.
Is function is unknown. Does Huntingtons disease provide a clue
to the function of huntingtin?
Probably not, which is the reason I indicated that Huntingtons disease is probably not a good example to illustrate pathways from gene to mind. Although the evidence is still incomplete, it looks as though huntingtin proteins that have extra long glutamine terminals tend to aggregate with one another and possibly with other proteins. This has been demonstrated in vitro by attaching glutamine tracts of different lengths to green fluorescent protein (GFP). In normal brains huntingtin is distributed throughout neurons, perhaps being enriched in nerve terminals, but also being observed in dendrites and cell bodies (and as stated before it is in non-neural cells as well). However, in brains from people with Huntingtons disease, and most clearly in mice expressing a transgene consisting of exon 1 of human huntingtin with an expanded CAG repeat, the protein (or the N-terminal fragment) is found tightly clustered within the nucleus in close association with ubiquitin and possibly with other proteins. At first it was thought that these inclusions were causing cell death, but other work suggests not. In cultured rat striatal neurons, mutant Huntingtin induced degeneration in cultured striatal neurons but not in hippocampal neurons. Neurons degenerated as a result of apoptosis, and cell death could be prevented by anti-apoptotic compounds and neurotrophic factors. In these experiments, huntingtin-induced neural degeneration was not correlated with intranuclear inclusions (Saudou et al, 1998). It will be fascinating to see how this story develops, but it is highly unlikely that we will find any special role for huntingtin in motor behavior or in cognition-- in the sense that the color pigments play a special and essential role in color vision. The process of DNA replication has been harnessed to allow the simple amplification of huge amounts of DNA. If a piece of single stranded DNA is placed in a mixture containing nucleotides, DNA polymerase and appropriate ions, the strand can serve as a template for the rapid synthesis of another strand of DNA if a short piece of complementary DNA is also present that will anneal to the single strand and prime the reaction. Kary Mullis had the brilliant insight that this processes could be harnessed by subjecting a mixture of DNA, primers and nucleotides to alternating cycles of temperature. A high temperature (close to the boiling point of water) causes the DNA to separate into single strands--a process called denaturation. The temperature is then dropped to allow the primers (short pieces of DNA, usually 20 nucleotides or so, that are complementary to the template) to anneal to the template, and the temperature is then raised somewhat to provide optimal conditions for incorporation of nucleotides into the growing strand (extension). Each time this cycle is repeated, the amount of DNA between the primers is in theory doubled. Thus, after 30 cycles, 1 ng of DNA could produce a gram of DNA. That much DNA is not required for most purposes, and the process is usually limited by the amounts of ingredients added. This process, called the polymerase chain reaction or PCR, has had a spectacular effect not only on biology, but also on such areas as forensics (it is this method that allows the DNA in a tiny spot of blood to be amplified and then typed, leading to unequivocal exclusion or matching with a known sample of DNA). For a fuller account of PCR, consult Genentech's PCR site. The following diagram is from Genentech's Access Excellence Site.  As an aside, Kary Mullis is not a typical Nobel Laureate. The following is an excerpt from his autobiography in The Prix Nobel: "My mother's parents were close to me all during my childhood, and her father Albert stopped by to see me in a non-substantial form on his way out of this world in 1986. I was living in California. "Pop" died at 92 and wondering what was happening to me out in California, stopped by Kensington for a couple days. My house afforded a view of San Francisco and the Golden Gate Bridge. His visit was an odd experience. Not at all frightening. I have cultivated the curious things in life and found this one pleasant. "Pop" and I sat in the evenings in my kitchen and I told him about the contemporary California world while we drank beer. I drank his for him as it appeared that although he was very much there for me, he was not there at all for the beer." Many methods exist for detecting mutations in DNA. The one used in our laboratory is called single strand conformation polymorphism and heteroduplex analysis (SSCP/HD). The name is more difficult than the method. In brief, when DNA is denatured into single strands and is then run on a gel that allows it to renature, each strand curls up into a distinctive conformation (or set of conformations) that migrates at a defined rate. Surprisingly, a change in a single nucleotide causes the conformation to differ enough so that it can be detected as a shift in band migration. The method is particularly good at detecting carriers of mutations, because the mutated and normal strands migrate differently, and a portion of them anneal to one another to form an anomalously migrating band called a hetroduplex. The worlds population has exceeded 6 billion people and is growing at a rate of about 2 people every second (click here to watch). Even given the high fidelity of DNA replication, it is expected that disabling mutations exist in some person, somewhere, for every human autosomal gene. For some mutations the frequency is high in certain populations: for example about 1 in 25 Caucasians in the U. S. carry a mutation for cystic fibrosis. Recent estimates of mutation rates suggest that they are higher than previously supposed. On average, each person carries 4 new mutations in addition to those inherited from past generations, and about 1.6 of these are deleterious enough to be eventually eliminated by selection.[ref] Once the sequence of a gene is known the gene can be amplified with PCR or by growing it in bacteria. The gene can then be linked to a promoter and placed into a circular piece of DNA, which is then placed into cells by a variety of methods. These cells will then express the protein from the DNA, and in that way the function of the protein can be ascertained. A variant of the PCR method can be used to introduce any desired mutation into a gene, once the sequence of the gene is known. The mutated gene can then be studied as indicated above. A spectacular extension of this method is to mutate genes in mouse embryonic stem cells, and then use these cells to produce a population of mice in which some of the offspring are homozyous for the mutated gene. Stem cells are immortal, and each retains the ability to grow into a complete mouse when implanted into a properly prepared donor female. Stem cells can be modified by selectively deleting genes (gene knockouts), or adding genes (transgenics). For gene knockouts only one copy of each paired gene is eliminated, so the resulting cloned mice must be bred until mice are produced that are homozygous for the deleted gene. Such mice can then be studied to determine the consequences of this gene deletion. A catalog of gene knockout mice lists the knockout mice using a variety of criteria. For example, one (short) catalog lists gene knockouts that primarily affect the nervous system. This technology is advancing rapidly, and is already mature enough to have generated for-profit companies that will make a transgenicor knockout mouse to your specification. One example of such a companyi s DNX Transgenic Sciences. Partial knockout experiments can also produce behaviorally interesting results. For example, Beverly Koller and her colleagues produced mice in which NMDA receptor expresssion is partly suppressed, and report that they display a pattern of behavior that is reminiscent of human schizophrenia. (Moln et al., 1999). (Please read: Genomics: Journey to the Center of Biology) Progress in molecular biological methods has been matched by spectacular progress in computer power, in bioinformatics, and in machines that carry out many of the procedures. All of these advances have made possible the industrialization of molecular-genetic research. For example, Celera Corporation has amassed 300 high throughput sequencing machines and is capable of decoding 140 million units of DNA every 24 hours. This has made possible the new science of genomics, in which the entire genome of an organism is sequenced and all genes identified. The human genome consists of ~100,000 genes and ~3,000,000,000 paired nucleotides. Perhaps 95% of the DNA is non-coding, leaving perhaps 150,000,000 nucleotides for the genes, or 1500 nucleotides per gene. After subtracting non-coding regions, the average gene product would be predicted to be less than 500 amino acids in length.A vast project to sequence the entire human genome is now underway. The origins of the human genome project are usually traced to a 1985 meeting on human genome sequencing held by Robert Sinsheimer at the University of California, Santa Cruz, leading Charles DeLisi and David Smith to develop plans for a Human Genome Initiative sponsored by the Department of Energy. NIH funding for human genome research began in 1987. The same year, DOE recommended a 15-year effort to map and sequence the human genome and designated a set of specialized human genome centers. The U.S. Human Genome Project formally began in October of 1990. In 1991 a genetic linkage map of the entire human genome was published, based on polymerase chain reaction/Sequence-tagged sites (PCR/STS). On 17 November 1999 the 1 billionth nucleotide of the human genome was sequenced, and on Decmber 1, 1999 the completion of the first chromosome (22) was announced. Spinoffs include the sequencing of genomes of other organisms. The entire genomes of many microorganisms have been completely sequenced and the databases can be searched via the web. In addition the genomes of a eukaryote Saccharomyces cerevisiae (baker's yeast) and of a multicellular organism, the nematode Caenorhabditis elegans have also been sequenced completely, and are available to anyone who has access to the Internet.
You can search these databases for
any sequence of interest. The fruits of this achievement will continue
to flourish for years or decades to come. Yeast has ~6,000 genes, and C.
elegans has ~19,000 genes that probably including representatives of most
human gene families. You might think that genes in our brains would be
one set of genes not represented in these lower organisms, but in fact
a burgeoning area of research has linked a large set of genes involved
in synaptic transmission with similar genes involved in vesicular trafficking
within yeast, and insights from C. elegans should be even more useful.
How can we go from genes to mind? A common estimate is that perhaps 30% of all human genes are specific to the nervous system. Examples of such genes are those that specify rhodopsin and the visual pigments, genes that make ions channels needed for nerve impulses, and genes for myriad receptors, neurotransmitters and complex synaptic machinery. Everyone seems to be comfortable with the idea that a single mutation can change one of the visual pigments so that color vision is dramatically altered. Numerous other mutations are known to affect the retina, leading to a variety of types of color blindness, night blindness, or complete blindness. These findings are readily accepted, and can be supported by very strong evidence. Logic suggests that other mutations will affect central processes, with consequences for virtually any psychological property. This proposition is not so readily accepted. Many reasons contribute to such skepticism. Supporting evidence is nowhere near as strong as it is for mutations that affect the retina, and the precision with which retinal mutations can be linked to visual changes is unlikely to be matched for most mutations that affect more central processes. But there is more to it than that, and the residual resistance can be linked to the same kind of thinking that finds something demeaning in the concept that we are biological machines. We may grant machine status to the retina, but as one moves centrally, psychological defenses become more formidable. While it is true that no link has been established between the vast majority of genes and any psychological property, that can be expected to change rapidly, and the rate of change can be expected to accelerate explosively in the coming years. How we use this new information is a central issue for modern society. |


