Introduction to Planar Cell Polarity
Acquisition of morphological asymmetry is critical to the physiological function of virtually all cells. Epithelial cells are polarized and carry out specialized functions on their apical and basolateral surfaces. Often, they are also polarized along an axis perpendicular to the apical-basal axis, producing patterned arrays within the epithelium (referred to as Planar Cell Polarity [PCP]). One beautiful example is the hair cells of the vertebrate cochlea, which are polarized such that their ciliary bundles, asymmetrically displayed on their apical surfaces, are optimally oriented for auditory signal transduction. PCP in the fruitfly, Drosophila, is evident in the orientation of cellular extensions called trichomes (or hairs), in the orientation of the multicellular bristles, and in the chiral organization and directional rotation of cell clusters that constitute the ommatidia, or facets, of the adult compound eye.
Genetic and molecular analyses have led to the identification of the seven-pass transmembrane protein Frizzled (Fz) and the four-pass tramsmembrane protein Van Gogh (Vang; aka Strabismus) as critical components of the PCP signaling mechanism. Ultimately, one side of the cell becomes uniquely marked by the asymmetric accumulation of Frizzled protein, while the adjacent side of the neighboring cell becomes marked by Vang. 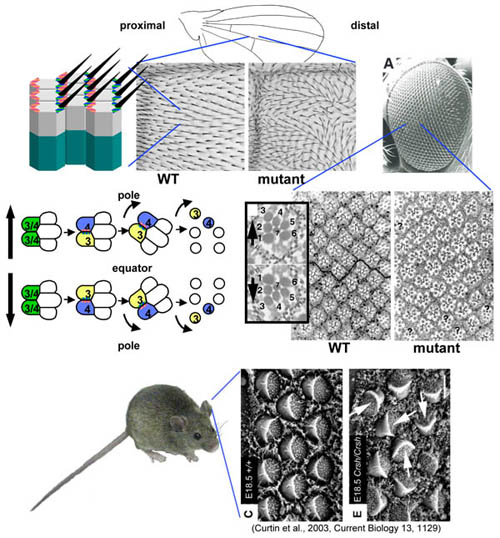
In the wing, Frizzled accumulates on the distal side of the cell, designating it as the future site for prehair formation, while Vang accumulates on the proxmial side. We have found that at the heart of PCP signaling, a core machinery mediates a competition between the proximal and distal proteins on adjacent surfaces of neighboring cells. This core machinery amplifies small differences to result in a highly asymmetric distribution of Frizzled and Vang. Other components of the core machine, including Dishevelled (Dsh; a conserved cytoplasmic protein), Prickle (a conserved, prenylated LIM domain protein) and Diego (Dgo) serve as components of a feedback loop that mediates the proximal-distal competition, and which themselves also become asymmetrically localized, either with (Dsh, Dgo) or opposite Fz (Vang and Pk). A consequence of this feedback loop function is that cells tend to align their polarity such that each cell accumulates high levels of Fz on the same side of the cell and high levels of Vang on the opposite side.
Layered on top of this local alignment mechanism, the PCP process requires a global directional cue to orient cell polarity with respect to the tissue axes. We believe that Dachsous and Fat (Ds and Ft; atypical cadherins) transduce a global cue that, through a microtubule based mechanism, biases the direction in which the feedback loop directs Fz accumulation, thus orienting Fz accumulation with respect to the tissue axes. Evidence indicates that inverted expression gradients of Four-jointed (Fj; a novel transmembrane protein) and Ds provide essential directional information in the eye, but that their graded expression is not essential, and can be replaced by uniform expression in the wing. Therefore, alternative modes of regulating Ds and Ft activity and providing directional information in the wing remain to be identified.
A third group of proteins have been shown by epistasis studies to function downstream of these other groups of proteins. Most of these show some tissue specificity, and while less is known about most of them, a reasonable view is that they couple signaling from the core proteins to the cell-type specific responses required to generate PCP in the individual tissues.
PCP is required for normal physiological functions of numerous adult tissues, as well as for the correct execution of developmental events. Hair cells in the mature cochlea are polarized to facilitate ciliary deflection in response to sound, and cells lining the reproductive and respiratory tracts have cilia whose polarized function is required for directed propulsion of luminal contents. PCP is implicated in controlling the patterns of hair growth on the skin of mammals, perhaps important for warmth and for efficient swimming. During development, neuroectodermal cells in the gastrulating vertebrate embryo polarize to facilitate convergent extension required for axis elongation and neural tube closure. In renal tubules, PCP appears to regulate both convergent extension and polarized cell divisions to control luminal diameter. PCP orients asymmetric cell divisions such that fate determinants are properly inherited by a single daughter (i.e. the Drosophila pI sensory organ precursor cells), PCP determines rotation and chirality of cell clusters (i.e. the Drosophila eye), and PCP determines the polarity of hairs on the fly cuticle, as described above.
Errors in PCP lead to human disease. Open neural tube defects can result from mutation of PCP genes in mice and humans. Disruption of PCP is associated with conotruncal heart defects and with heterotaxy. Some forms of ciliary dyskinesias appear to result from disruption of PCP in the respiratory airways. A human epilepsy/ataxia syndrome results from mutation of several PCP genes. Understanding how PCP signaling mechanisms contribute to these conditions may lead to improved diagnosis and treatments.