| |
Biology 137 2008 Spring Semester
Meta-report 4: Assessing the Effects of Metals on Dahlia Pigmentation

Dahlia 'Lakeview Premier', a white formal decorative type,
with flowers that can change color based on metal ion complexation with anthocyanins
Introduction
Petal coloration is important in attraction of pollinators to flowers. The diversity of flower color has been intensively studied and utilized in genetic research since Mendel. Anthocyanin pigments are of central importance in flower coloration in many angiosperms, including dahlias and other Composites, although pigment molecules alone are often insufficient to produce the colors seen in many flowering plants. Within a cell anthocyanin is synthesized in the cytoplasm, but only after transfer to the vacuole does the pigment attain a bright color. Among the accessory factors that determine coloration is the pH of vacuoles of floral cells, anthocyanin assembly with flavonols and flavones and other organic molecules (collectively called co-pigments) (Reuveni et al. 2001, Mol 1998), and anthocyanin assembly into complexes coordinating a metal ion such as aluminum and copper (Sarma et al. 1997) or magnesium and manganese (Shaked-Sachray et al. 2002). Some bright blue flowers coordinate a chromate ion with 4 anthocyanin molecules. Depending on the protonation state, anthocyanins will absorb different wavelengths of light; in acidic conditions, anthocyanins typically appear red while in basic conditions they are usually green or blue. For instance, in petunia, low vacuolar pH correlates with red coloration, whereas high pH correlates with purple (Reuveni et al. 2001). However, the availability of metals ions generated by pumps that selectively transport specific ions into the vacuole can also have an effect on flower pigment. Hydrangeas flowers are blue when grown in acidic soil and red when grown in alkaline conditions, which is contrary to what would be expected based on pH affecting anthocyanins. The mechanism which causes this color change was found to be pH affecting the solubility of aluminum ions; those aluminum ions in turn were responsible for floral pigmentation (Yoshida et al. 2008, Takeda et al. 1990, Takeda et al. 1985). To assess whether environmental pH or metal ion conditions might modulate flower color in dahlias, metal or pH treatments were applied to several Dahlia cultivars and the effects on flowers examined. Metals selected were:
This report synthesizes the analysis of the effects of metals ions and pH in different Dahlia cultivars conducted by students in the Spring 2008 quarter of Biological Sciences 137; Dahlias in Plant Genetics: E. Abrash, A. Christian, R. Kamber, J. McCallen, A. Schultink, W. White, and C. Young.
Objectives
The objectives of this study followed the outline of metals used above; analysis of which metals or pH affected dahlia floral pigments was the primary objective of this experiment, and results will be defined based on the individual researcher and the metals or pH values used.
Materials and Methods
In this investigation, groups of plants were dosed with the various metal treatments in differing concentrations. Doses were applied twice during a week to plants in single watering trays (for each treatment) such that subsequent watering would promote uptake of additional metal or hydrogen ions. In each case, about 125 mL of total solution was given to each tray over seven days. Alternately, for plants in individual pots, metal solutions were applied on the first day and the fourth day of the experiment. Applications of acids were typically in the pH range of 4-5, through addition of citric acid. Application of bases were in the 8-9 pH range through addition of sodium hydroxide. Copper, iron, magnesium, manganese, molybdenum and cobalt were given using 10 μM solutions. Zinc was dosed using 1 μM and 5 μM solutions as separate treatments. Tin and nickel were given using 50 μM solutions.
Photographs of flowers were taken at the time of initial treatment. One week later, plants were re-examined for changes in flower color and compared to individuals of the same cultivar that had not received the treatments, and compared to ADS color chips.
Summary Table of Results
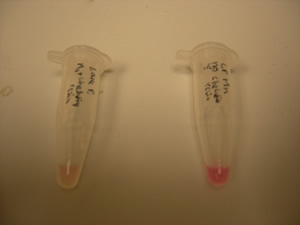
Figure 1. Dahlia Lakeview Premier changes color in the presence of magnesium (right) and manganese (left) when compared with control (center).
|
|
|
|
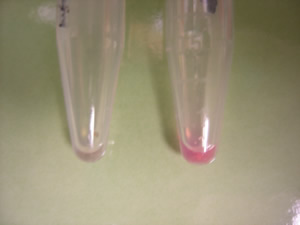
Figure 2. Dahlia Moray Susan changes color in the presence of cobalt at 10 uM (right) and cobalt at 20 uM (left) when compared with control (center).
|
|
|
|
Figure 3. Dahlia Dixie's Winedot failed to show color change in the the presence of copper (right) and citric acid (left) when compared with control (center), although treatments did accelerate senescence.
|
|
|
|
Results and Discussion
Plants in most treatment groups displayed very little pigmentation differences compared to control plants. Instead, flowers on most treated cultivars appeared to senesce prematurely, displaying a dry and crinkled appearance despite apparently adequate health of the vegetative tissues. Certain treatments exacerbated this effect: copper in particular as well as zinc and basic solutions seemed to cause premature senescence. The observed senescence phenotypes might reflect a specific effect of the treatments on floral progression; alternatively, floral senescence might be symptomatic of an overall reduction in plant health, with flowers being sensitive “indicator” organs that display phenotypes before leaves and other tissues. On the other hand, better control of flower age might help to demonstrate the effects of metals on senescence, as some of the control flowers aged significantly during the duration of the experiment. Additionally, natural variation in senescence phenotypes, which would be especially prevalent in the vastly differentiated floral forms of dahlias, has previously been observed in Arabidopsis thaliana (Levy et al. 2005).The experimental observations suggest that at the concentrations used, only magnesium, manganese, and to a lesser extent cobalt affect flower color.
Cobalt treatment followed three phases of pigmentation. The flowers of the cultivar Moray Susan before treatment were pink with yellow coloring in the interior. About a week into treatment the flowers minimized their yellow color but the pink became more red, resembling a magenta pigment. After extensive cobalt treatment, the flowers regained their pink color, but only in the outer ray florets. The inner ray florets did not show any pink but were completely yellow.
Lakeview Premier plants receiving magnesium and manganese displayed significant differences in ray floret coloration. Both magnesium and manganese have been shown in the past to complex with anthocyanin pigments (Takeda and Hayashi 1977, Mazza and Miniati 1993). Magnesium has furthermore been shown in a recent study to stabilize anthocyanin pigments in flower petals, not by increasing anthocyanin synthesis, but most likely by forming stable anthocyanin/magnesium complexes (Shaked-Sachray et al. 2002). If manganese and magnesium from the watering solution are translocated to developing flowers, it seems plausible that anthocyanin synthesis and/or accumulation would be upregulated in the presence of Mg 2+ or Mn 2+. Magnesium has shown to be a component of the blue pigment commelinin from cornflowers (Hayashi and Takeda 1970) and manganese toxicity has been correlated with increased anthocyanin accumulation in spearmint (Asrar et al. 2005). Definitive determination of the presence of manganese and magnesium in darkened flowers should be established. If present, the nature of the metal-anthocyanin interaction(s) could also be more fully investigated. Of particular interest would be identification of the anthocyanin pigments present in Lakeview Premier through UV-visible spectrophotometry.
Certainly the results shown here represent only an initial step in investigating confounding effects of anthocyanin pigmentation. Further research might help determine exactly which anthocyanins are present, their degree of complexation to metal ions, and the effects of ranges of metal ion concentrations and pH levels. Could we treat at a lower dose of metal for a longer period of development and see more changes in flower color and avoid the senescence response?
Figure 4. Dowex chelating resin can extract metal and alter anthocyanins from dark purple to light pink. |
|
|
|
One experiment performed in the lab was to extract anthocyanin in acidic methanol (1 mL HCl in 100 mL in methanol) from several pink and purple cultivars. The extracts were bright pink or purple. When extracts were mixed with Dowex chelating resin, anthocyanins were converted to a pale yellow-pink color. Dowex can extract divalent metal ions (depending on the affinity of the pigment for the metal). Adding back metals could restore some color to the anthocyanin in the pink-purple range; adding ferrous iron caused the anthocyanin to be reddish brown. In the future, it would be interesting to explore this further by identifying which ions are bound to Dowex from the extract. Do all dahlias complex the same metal ions? Another interesting question would be could we treat at a lower dose of metal for a longer period of development and see more changes in flower color?
References
Asrar, Z., Khavari-Nejad, R. and H. Heidari. 2005. Excess manganese effects on pigments of Mentha spicata at flowering stage. Archives of Agronomy and Soil Science. 51: 101-107.
Brenchley, W.E. 1938. Comparative effects of cobalt, nickel and copper on plant growth. Annals of Applied Biology. 25: 671-694.
Cohen, B.B. 1940. Some effects of stannous sulphate and stannic chloride on several herbaceous plants. Plant Physiology. 15: 755-760.
Hayashi, K. and Takeda, K. 1970. Further purification and component analysis of commelinin showing the presence of magnesium in this blue complex molecule. Proceedings of the Japan Academy 46: 535-540.
Levy, S., and A. Wingler. 2005. Natural variation in the regulation of leaf senescence and relation to other traits in Arabidopsis. Plant, Cell and Environment. 28: 223-31.
Mazza, G., and E. Miniati. 1993. Anthocyanins in fruits, vegetables, and grains. CRC Press, Boca Raton.
Mol, J., Grotewold, E., and R. Koes. 1998. How genes paint flowers and seeds. Trends in Plant Science. 6:
212-217.
Reuveni, M., Evenor, D., and B. Artzi. 2001. Decrease in vacuolar pH during petunia flower opening is reflected in the activity of tonoplast H+-ATPase. Journal of Plant Physiology. 158: 991-8.
Sagasser, M., Lu, G.H., Hahlbrock, K., and B. Weisshaar. 2002. A. thaliana TRANSPARENT TESTA 1 is involved in seed coat development and defines the WIP subfamily of plant zinc finger proteins. Genes and Development. 16: 138-149.
Sarma, A., Sreelakshmi, Y. and R. Sharma. 1997. Antioxidant ability of anthocyanins against ascorbic acid oxidation. Phytochemistry. 45: 671-674.
Shaked-Sachray, L., Weiss, D. Reuveni, M., Nissim-Levi, A., and M. Oren-Shamir. 2002. Increased anthocyanin accumulation in aster flowers at elevated temperatures due to magnesium treatment. Physiologia Plantarum. 114: 559-565.
Takeda, K., Yamashita, T., Takahashi, A., and C.F. Timberlake. 1990. Stable blue complexes of anthocyanin-
aluminium-3-p-coumaroyl- or 3-caffeoyl-quinic acid involved in the blueing of Hydrangea flower. Phytochemistry. 29: 1089-1091. 1990.
Takeda, K., Kariuda, M., and H. Itoi. 1985. Blueing of sepal colour of Hydrangea macrophylla. Phytochemistry. 24: 2251-2254.
Takeda, K. and K. Hayashi. 1977. 1. Metallo-anthocyanins: Reconstruction of commelin from its components, awobannin, flavocommelin and magnesium. Proceedings of the Japan Academy. 53: ser. B.
Tso, T.C., Sorokin, T.P., and M.E. Engelhaupt. 1972. Effects of some rare elements on nicotine content of the tobacco plant. Plant Physiology. 51: 805-806.
Yoshidaa, K. et al. 2008. Change of color and components in sepals of chameleon hydrangea during maturation and senescence. Nagoya University, Japan.
Download the Text (.doc)
        
|