| |
Biology 137 2008 Spring Semester
Meta-report 5: Flavonoid Identification and Genetic Analysis of Variegation in Dahlia Dixie’s Winedot

Dahlia 'Dixie's Winedot'', a red and yellow variegated informal decorative type,
where variegation presumably occurs through transposon activity disruption of the anthocyanin pathway
Introduction
Dahlias have been developed through classic breeding, but with modern genetics scientists are starting to gain a deeper understanding of what contributes to flower coloration and how it can be manipulated. This study sought to explore the mechanism behind the variegated flower coloration of the Dahlia cultivar Dixie’s Winedot.
Dixie’s Winedot flowers have a pale yellow background color intermixed with dark streaks of purple, hence the classification as a variegated variety. There are several other variegated varieties of Dahlia, which typically follow the pattern of having a pale background color with darkly pigment streaks. The streaks can range in size from just a thin strip to large sectors or even an entire flower. The streaks are typically radial from the center of the flower. The pale background color and the streak distribution suggest that the variegation is the result of transposon activity (Chawla 2000).
In the transposon model for variegation, the plant is homozygous for non-functional recessive alleles for one step in the pigment pathway. One (or more) copies of the nonfunctional alleles contain a transposon, which is disrupting the functionality of the gene. This transposon is capable of spontaneously excising from the gene (under the control of a transposase protein); if after the transposon excises the host repairs the DNA to restore the correct open reading frame, the functionality of the allele is restored and the pigment pathway is restored. The resulting purple sector is called “revertant” and the restored allele is a revertant allele. The restored gene is passed on to subsequent daughter cells during mitosis and cell division. The daughter cells are typically arranged in a line radially from the center of the flower reflecting the patter of cellular division in the developing petal tissue: there are more cell divisions to increase petal length than width. This creates a streak of cells that have the functional gene and hence have a darker pigmented color. If the transposon excises very early in flower development, then a larger sector will likely develop because the gene will be passed on to more daughter cells. Smaller sectors and streaks are caused by excision of the transposon later in petal development; excisions that occur after the final cell divisions produce a single dark cell. Previous studies of similar variegations in other species have found this model to be accurate (Habu et al. 1998). This information can be used to construct an analysis of petal (or flower) development by observing the shape and frequency of dark colored sectors, i.e. observing a 1/2 flower means that at some time there were just 2 cells responsible for making the flower.
It was hypothesized that a single gene in the anthocyanin biosynthetic pathway is disrupted by a transposon in normal flowers of Dixie’s Winedot (Ogata et al. 2004). This experiment set out to confirm that transposon activity was responsible for the variegated phenotype by blocking or activating a step in the flavonoid pathway (Figure 1). A full revertant plant first identified as a branch with purple flowers on a variegated plant; cuttings from this branch were rooted to produce a fully purple flowered plant to use to the variegated original stock. (The original stock plant will be referred to as “wild type” in this paper; student use of “wild type” should in fact refer to the wild type of Dahlia sp., as found naturally occuring in northern Central America.)
Largely conserved throughout the plant kingdom, flavonoids serve multiple functions in most plant families , including protection from mutagenic UV rays, pollinator-attraction, feeding deterrance and microbe defense (Mol et al. 1998, Winkel-Shirley 2001, Winkel-Shirley 2002). The flavonoid biosynthetic pathway begins with chalcone-isomierized condensation of one molecule of 4-coumaroyl-CoA and three molecules of 3-malonyl-CoA to yield chalcone. Chalcone is subsequently isomerized to naringenin by chalcone isomerase, at which point it can either be maintained as such or shuttled into several different pathways to give rise to six different classes of flavonoids: the chalcones, flavones, flavonols, flavandiols, anthocyanins, and condensed tannins (Mabry et al. 1970, Markham 1982, Winkel-Shirley 2001).
This report synthesizes the analysis of the control of variegation in Dixie’s Winedot conducted by students in the Spring 2008 quarter of Biology 137; Dahlias in Plant Genetics: A. Schultink and C. Young.
Objectives
The objectives of this study in Dixie’s Winedot were:
-
Identify the pigment accumulating in recessive (yellow) cells. It was assumed that the flavonoid product accumulating would be the one before the step requiring the nonfunctional gene. By identifying the accumulated yellow pigment and knowing the biosynthetic pathway, the nonfunctional gene could be identified. The pale yellow background color is typical of blocks very early in the pathway, and it was hypothesized that the accumulating pigment was a chalcone. To determine if this was indeed the case, thin layer chromatography (TLC) was used to separate pigments and UV spectrophotometry was performed to identify the accumulating flavonoid.
-
Use polymerase chain reaction ( PCR) ampflication with a primer designed for chalcone isomerase (CHI), the presumed block in the pathway. If there is a transposon in the “variegated” gene, the PCR would yield either no product (transposons often block the PCR reaction) or a product much larger than in the revertant plant; with the transposon excised, the revertant gene should be smaller.
Materials and Methods
Flavonoid Analysis
Petals of Dixie’s Winedot were ground and extracted with 2 mL of acidified methanol (100 mL methanol and 1 mL 1M hydrochloric acid) from both the wild type and the revertant. Both of these extracts looked similar. To facilitate pigment identification, TLC was performed using microcrystalline cellulose plates and acetic acid, hydrochloric acid, and water solutions. Bands of interest where scraped from the TLC plate and the pigment(s) was resuspended in acidic methanol. The absorbance spectrum of the extracts was taken between 250 and 600 nm. Standards were prepared and compared to the unknowns using TLC and UV-visible spectrophotometry.
Genetic Analysis
Primers were designed for the dahlia CHI gene by first comparing CHI gene sequences from several plants as to date there is no gene information from Dahlia. Asteraceae sequences of CHI mRNA from sunflower, two types of Chrysanthemum, Callistephus chinensis, and Saussurea medusa have been sequenced and were used. An alignment was performed on these sequences using ClustalX2, and PriFi; once aligned, regions of high conservation within the Asteraceae were identified and these regions were subsequently used to design suitable primers for CHI. Primers were selected in highly conserved regions of the gene and to cover the majority of the gene. This allowed for the design of primers that should allow amplification of the dahlia gene (Figure 5). Because the sequences did not match up perfectly there was some degeneracy in the primer designs, because prior to sequencing the dahlia gene there is no way to predict the precise sequence of the dahlia gene. The primers for primer pair 1 were:
- Primer pair one Forward 5'- CCTMGGWGGCGCMGGTGTGAGAGGTATGG
- Primer pair one Reverse 5'-TTGTRAAGAGAATAGAKGMGCCTGG
- Primer pair two Forward 5'-TAACNGGTAAGCAATACKCTGA
- Primer pair two Reverse 5'-AGTCTTGAWGCCAAACTTTGTTT
The degeneracy is indicated by the non ATCG base pairs listed. The melting temperatures predicted for these four primers were 71ºC, 58ºC, 60ºC, and 61ºC, respectively. To perform PCR, dahlia DNA was extracted and purified using a CTAB extraction (following protocol of Timmerman-Vaughan et al. 2005). A high performance Taq polymerase was used for amplification of the extracted DNA; this allowed for amplification of even very large fragments. Because the optimal annealing temperatures for the primers were unknown, 12 PCR reactions were run for each primer pair across a gradient from 50 to 60 ºC. Higher temperature allowed more specific amplification while lower temperature ensured strong annealing. The PCR products were separated by size using electrophoresis on a 1.2% agarose gel at 100 V for 45 minutes. Five microliters of PCR product was used with 5 uL of loading dye in each lane of the gel. Poor resolution resulted from the user-prepared gel, therefore the gel eletrophoresis was repeated with a commercial E-Gel.
Figure 2. Primer selected for use in this experiment. Six known cDNA sequences for CHI from the Asteraceae family were used to engineer primers using multisequence primer selection tool PriFi. Two primer pairs were selected. The known introns from the petunia CHI-B gene are shown. |
|
Results and Discussion
Flavonoid Analysis
Based on TLC (Figure 3), differences between the Dixie’s Winedot variegated yellow with purple streaks and the fully purple revertant extracts were clearly visible. Two bands clearly stood out as different between the two samples: band 1 was yellow in color and much more prevalent in the wild type; band 2 was pink / purple in color and much more prevalent in the revertant.
Figure 3. A solution of acetic acid, HCl and water was used to separate the wild type and revertant extracts on a microcrystaline cellulose plate using TLC. |
|
Figure 4. The spectra for each of the two bands isolated from the samples are shown above. Band 1 was much more concentrated in the wild type whereas band two was more concentrated in the revertant. The band 2 spectrum, with a peak around 525 nm, is characteristic of an anthocyanin. Note: in this figure WT (wild-type) is used to refer to the original Dahlia Dixie’s Winedot stock. |
|
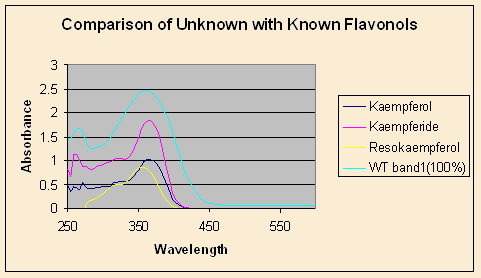
Figure 5. WT Band 1 compared to known standards Kaempferol, Kaempferide and Resokaempferol. |
|
An identification of the unknown band 1 area was attempted using UV-visible spectroscopy of standards (Figure 4). The unknown spectrum did not match up well with that of chalcone, because it contained a peak around 270 nm that chalcone lacks (Figure 5). The peaks of the unknown at 270 nm and 362 nm suggest that the unknown is a flavonol rather than a chalcone. There are several flavonols that are part of the anthocyanin pathway, and flavonols can accumulate as “side reaction products” when the anthocyanin pathway is blocked. The unknown was compared to the spectra of three flavonols: kaempferol, resokaempferol, and kaemferide. The spectrum of the compound isolated from band 1 appears to be similar to that of kaempherol or some other flavonol, with major peaks near 360 and 270 nm. From the model proposed, these flavonols are not the products expected to be accumulating. The stability of flavonols would favor accumulation if the pathway was blocked at the DFR gene. Nonetheless, other studies have shown that this is generally not the case: analysis of Petunia hybrida mutants lacking DFR and F3’H activity showed that dihydrokaempherol, not kaempherol, accumulated (Linn et al. 1990). Additionally, the unknown spectrum lacks the characteristic peak around 325 nm that flavonols exhibit. Therefore, while some flavonols may be present in the unknown from band 1, the main yellow pigment is probably not a flavonol.
From the pathway shown in Figure 1 and the hypothesis that CHI was the defective gene, the accumulating product was expected to be chalcone, but the unknown spectrum does not match up well with that of chalcone. It is possible that the accumulation of chalcone led to the synthesis of some other flavonoid. Defective CHI has been shown in the previous studies to lead to the accumulation of isosalipurposide as well as chalcone (Froemel et al. 1985, Reuber et al. 1997). The broad peak of the unknown matches up well with that of isosalipurposide obtained from the literature. Also, chalcone is known undergo a slow isomerization to the racemic flavanone. One of these enatiomers is narigenin, which can continue the anthocyanin pathway. It is possible that a defective CHI may lead to the accumulation of isosalipurposide, chalcone, and small amounts of the rest of the pigments in the pathway, including the flavanones and flavanols. The tentative conclusion is that the unknown spectrum is resultant of such a mixture, however more work remains to confirm this hypothesis. HPLC analysis of the isolated unknown from the band 1 area would likely yield more information about the number of pigments types present (Reuber et al. 1997). Furthermore, these pigments could be collected and analyzed by UV-visible spectroscopy individually; by viewing the pigments in isolation a more definitive identification could be made.
Genetic Analysis
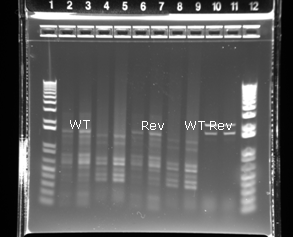
The gel of all the PCR products is shown in Figure 6. The results of primer pair one are difficult to interpret because the image is saturated. For primer pair two, two discrete fragments are visible at higher temperatures, while more bands are visible at lower annealing temperatures. A second gel was run using a subset of the PCR products. This gel provided better resolution than the first and allowed for better size estimation of the fragments (Figure 7). The two fragments from primer pair 2 appear to be about 1.7 and 2.1 kb. Numerous fragments are visible for primer pair one, ranging from 350 bp to 1.6 kb.
Figure 6. PCR products were visualized using gel electrophoresis. A temperature gradient from 50ºC to 60ºC was used for the annealing temperature during the PCR. |
|
Figure 7. An E-Gel was run to obtain better resolution of the PCR products. The primer one products from WT and revertant were compared from high and low temperature in lanes 2-9. One ul and two ul of PCR product were ran in neighboring lanes for primer pair one. |
|
Primer pair one seemed to result in significant non-specific amplification. A higher annealing temperature may improve the specificity, particularly because the melting temperature for the forward primer was predicted to be 71ºC. One concern is intron location, which can only be inferred from gene sequences of other plants, because the gene has not been sequenced from Dahlia. If the petunia intron locations are representative of those in Dahlia, primer pair one is likely to be ineffective. The intron positions and sizes for Petunia hybrida CHI-B are shown in Figure 2. The CHI protein sequences for Petunia and Asteraceae were shown to be similar, so such a comparison is valid. The intron predicted in the forward primer of primer pair one would prevent annealing and successful amplification of the desired sequence.
Primer pair two was more successful at achieving specific amplification. It is likely that these fragments are the result of the amplification of the desired gene sequence. The fragments were sized at about 2.1 and 1.7 kb. This is significantly larger than the expected band of 0.5 kb. This could be explained by the dahlia gene having larger introns than the petunia gene. Primer pair two resulted in the amplification of two fragment lengths. There are several possible explanations for this: dahlias could have two different CHI genes of different sizes (Petunia is known to have two different CHI genes, CHI-A and CHI-B), and amplification of these different genes could explain why two different fragment lengths were produced; alternately, there may be two different alleles of the same gene present that have different sizes. If the latter explanation is correct, the different sizes of these alleles could be explained by one of the alleles having a 0.4 kb transposon inserted into it; because the variegated flower has streaks on purple, some of this allele should be present in the wild type sample. Even though the revertant flower is entirely purple, if the wild type is homozygous for the transposon allele then the revertant plant is heterozygous: one copy of the revertant allele and one copy of the transposon-carrying allele. The results of the PCR with primer pair two are consistent with this hypothesis but they do not rule out other explanations. Future work could be conducted to provide more evidence to either support or discredit the hypothesis proposed. For example, the PCR products from primer pair two could be sequenced. If the two sequences were identical except for a transposon-like insert this would be strong evidence that the hypothesis was correct. Alternatively, the PCR with primer pair two could be repeated on other Dahlia varieties without variegation to see if the two band result is specific to Dixie’s Winedot. Even if the was not a transposon present in the area of the gene covered by primer pair two, there could be a transposon located somewhere else in the gene.
Ultimately the full DNA sequence of all of the anthocyanin pathway genes of Dahlia would be highly desirable so that allelic variation among Dahlia varieties could be examined in detail and correlated with pigment shades and intensities. The DNA sequence knowledge could also be used to discover and classify the types of transposons generating variegation in the many Dahlia varieties with this type of pigmentation pattern.
References
Chawla H.S. 2000. Introduction to Plant Biotechnology. Enfield: Science Publishers.
Froemel, S., de Vlaming, P., Stotz, G., Wiering, H., Forkmann, G., and A.W. Schram. 1985. Genetic and biochemical studies on the conversion of flavanones to dihydroflavonols in flowers of Petunia hybrida. Theoretical Applied Genetics. 70: 561-568.
Habu, Y., Hisatomi, Y., and S. Lida. 1998. Molecular characterization of the mutable flaked allele for flower variegation in the common morning glory. The Plant Journal. 16: 371-376.
Linn, F., Heidmann, I., Saedler, H., and P. Meyer. 1990. Epigenetic changes in the expression of the maize A1 gene in Petunia hybrida: role of numbers of integrated gene copies and state of methylation. Molecular Genetics and Genomics. 222: 329-336.
Mabry, T.J., Markham, K.R., and M.B. Thomas. 1970. The systematic identification of flavonoids. Journal of Molecular Structure. 10: 320.
Markham, K.R. 1982. Techniques of Flavonoid Identification. New York: Academic Press.
Mol, J., Grotewold, E., and R. Koes. 1998. How genes paint flowers and seeds. Trends in Plant Science. 3:
212-217.
Ogata, J., Sakamoto, T., Yamaguchi, M., Kawanobu, S., and K. Yoshitama. 2004. Isolation and characterization of anthocyanin 5-O-glucosyltransferase from flowers of Dahlia variabilis. Journal of Plant Physiology. 158: 709-714.
Reuber, S., Jende-Strid, B., Wray, V., and G. Weissenbock. 1997. Accumulation of chalcone isosalipurposide in primary leaves of barley flavonoid mutants indicates a defective chalcone isomerase. Physiologia Plantarum. 101: 827-832.
Timmerman-Vaughan, G.M., Pither-Joyce, M., and G. Clover. 2008. A positive-control PCR assay that is based on chloroplast ndhB sequences and applicable to nucleic acid extractions from diverse plant taxa. Plant Molecular Biology Reporter. 23: 77-78.
Winkel-Shirley, B. 2002. Biosynthesis of flavonoids and effects of stress. Current Opinion in Plant Biology. 5: 218-223.
Winkel-Shirley, B. 2001. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiology. 126: 485-493.
Download the Text (.doc)
         
|